Photosynthesis: Turning Sunlight, Water and Air into Pecans

Figure 1. The relationships of photosynthesis, stomatal conductance and transpiration to plant water status (given as midday stem water potential) in 'Western' pecan (from Othman et al., 2017)
Green-colored pigments—chlorophyll a and chlorophyll b—are found in the chloroplasts of leaf cells. Inside the chloroplasts, the leaf cells capture and convert the energy from blue and red (but not green) wavelengths of sunlight into chemical energy through a series of chemical reactions, called light reactions. Like batteries that store portable energy, two phosphorus-containing molecules abbreviated as NADPH and ATP hold this chemical energy. This chemical energy is more useful for the plant than raw light energy because it can be safely stored for a short time and transported within the chloroplast.
The chemical energy from the light reactions is moved along into another set of chemical reactions in the chloroplast called the dark reactions, or Calvin Cycle. The term “dark reactions” is misleading because the dark reactions in most plants (including pecan) do not actually work in the dark. There are three reasons for this: 1) the dark reactions depend on NADPH and ATP from the light reactions, and when the lights go out, the light reactions also stop; 2) the dark reactions are catalyzed by some enzymes that the dark deactivates; and 3) the stomata (pores on the leaf’s underside) have guard cells that are sensitive to light, closing like a gate in the dark. Closed guard cells block carbon dioxide from entering the leaf through the stomata, and as a result, the dark reactions slow to a stop.
In the dark reactions, the NADPH and ATP chemical energy from the light reactions along with carbon dioxide and water are used to build carbohydrates, which can be thought of as a different form of chemical energy. These first simple three-carbon sugars are converted into a wide range of other larger carbohydrates that have various uses by the tree.
Some of the sugars are made into glucose, a 6-carbon sugar, that may be readily used by cells for metabolism. Just like in your cells, the energy from the glucose is used or burned to supply cellular respiration so that all of the cellular processes can proceed normally. The byproduct of this, just like in your cells, is carbon dioxide gas. (Photosynthesis rates are usually much higher during the day than respiration rates so that there is a net uptake of CO2 by the plant; at night, the situation is reversed.)
On the other hand, if the glucose is not used up in cellular respiration, the plant can use the carbon from the glucose to manufacture all of the building blocks that make up the plant: cellulose for cell walls, lignin for the woody tissues (the branches, trunk, and nutshells), lipids for the kernel oils, and so on. Really every part of a pecan tree is made mostly from carbon that was fixed in photosynthesis. Again, similarly as with a growing human (child or adult!), a pecan tree grows and gains weight only by incorporating the carbon from its food. The difference here is that the pecan tree makes its own food.
https://www.canva.com/design/DADoesOMEbk/view
Not all of the products of photosynthesis are necessarily used by the tree right away. Some of the sugars from photosynthesis are converted to sucrose, a 12-carbon disaccharide sugar, which the tree uses to transport food around the plant in the phloem tubes (i.e., in the bark) to the various parts of the tree that need it. At times, the sucrose may have to travel very, very long distances from the photosynthesizing leaves, such as growing root tissues. Finally, some of the sugars from photosynthesis can be made into starch, which plant cells can store for relatively short (overnight) or much longer (over the winter) periods when photosynthesis isn’t happening. Starch is one of the major forms that pecan trees store their dormant season food reserves when there is not any current photosynthesis.
The water molecule is a raw ingredient of photosynthesis, but water is also essential for photosynthesis for another reason. The water status of a plant—how thirsty it is—affects its leaf stomatal conductance. The stomata are the pores on the bottom of pecan leaves that allow carbon dioxide to enter the air spaces inside the leaf tissues where it may diffuse into the cells where the dark reactions of photosynthesis take place. For well-watered plants, the guard cells that surround each stomatal pore-like gates will typically open up, allowing maximum potential photosynthesis to occur. But as a plant becomes thirsty, the guard cells on the stomata close, blocking carbon dioxide from entering the air spaces inside the leaf tissues. When the stomata close, photosynthesis declines even when the light levels are high (Figure 1).
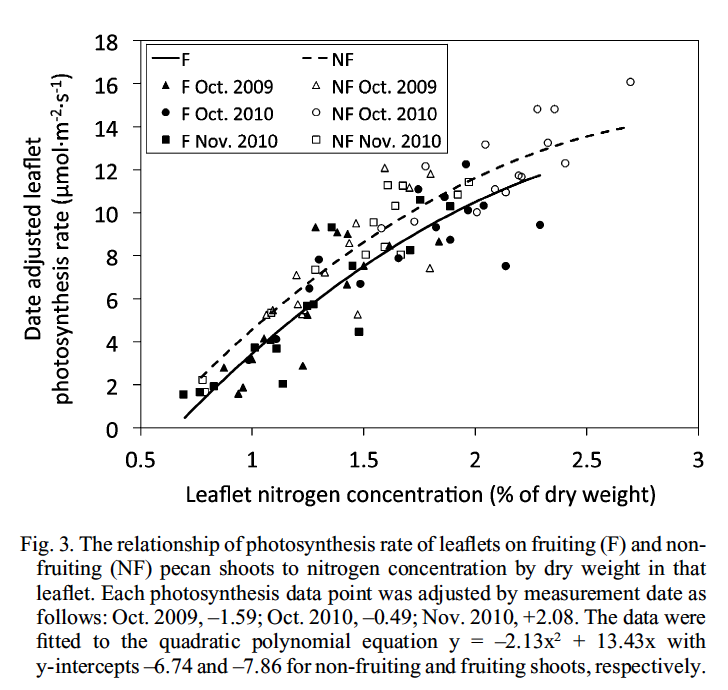
Figure 2. The relationship between photosynthesis rate and leaf tissue nitrogen concentration in ‘Western’ pecan (from Heerema et al., 2014)
To conduct photosynthesis, pecan trees also need certain mineral nutrients. These nutrients are not really “ingredients” for the products of photosynthesis (except for phosphorus, which is a kind of “ingredient” in some types of sugars—but that’s a different story) but rather, necessary components. Nitrogen is one of these nutrients. Nitrogen can be thought of as part of the machinery that runs just about everything in the light and dark reactions of photosynthesis because it is part of every one of the enzymes involved in these chemical reactions, the chlorophyll molecular structures, ATP and NADPH, and so on. In fact, during the growing season, more nitrogen may be found in just one enzyme of the dark reactions (and enzyme nicknamed Rubisco) than in any other molecule in the whole plant! It should come as no surprise then that when nitrogen levels are low in leaf tissue, leaf photosynthetic rates are also invariably low (Figure 2).
Another mineral nutrient important for photosynthesis, magnesium is a part of the molecular structure of chlorophyll. Phosphorus is part of the structure of many of the biochemicals that hold chemical energy (including ATP and NADPH). Some of the micronutrients, including iron, manganese, and zinc, are cofactors for enzymes involved in various ways with photosynthesis.
Pecan growers cannot control everything in their orchards that might affect photosynthesis. They do not have much control over the ambient carbon dioxide concentrations or the air temperature, both of which can affect photosynthesis rates. However, they have a great deal of control over certain other factors. Growers can control the tree’s water and nutrient status through optimizing their irrigation and fertilizer programs. More than that, pecan growers can work to keep photosynthesis high in their orchards by managing certain insect pests. Black pecan aphid feeding causes chlorotic and necrotic spots to appear on leaves, which reduces the effective photosynthetic leaf area in the canopy. On the other hand, the black margined aphid produces copious amounts of honeydew that creates a substrate for sooty mold growth on the leaf surface, and thus, blocks sunlight capture. While planning management practices, growers should think about photosynthesis and which affecting factors can be managed to ensure their trees are getting all the support they need to thrive.
