Pecan Aphids, Part II: Feeding Biology
In this second part of a three-part series, Entomologist Ted Cottrell dives into the black pecan aphid's feeding practices.
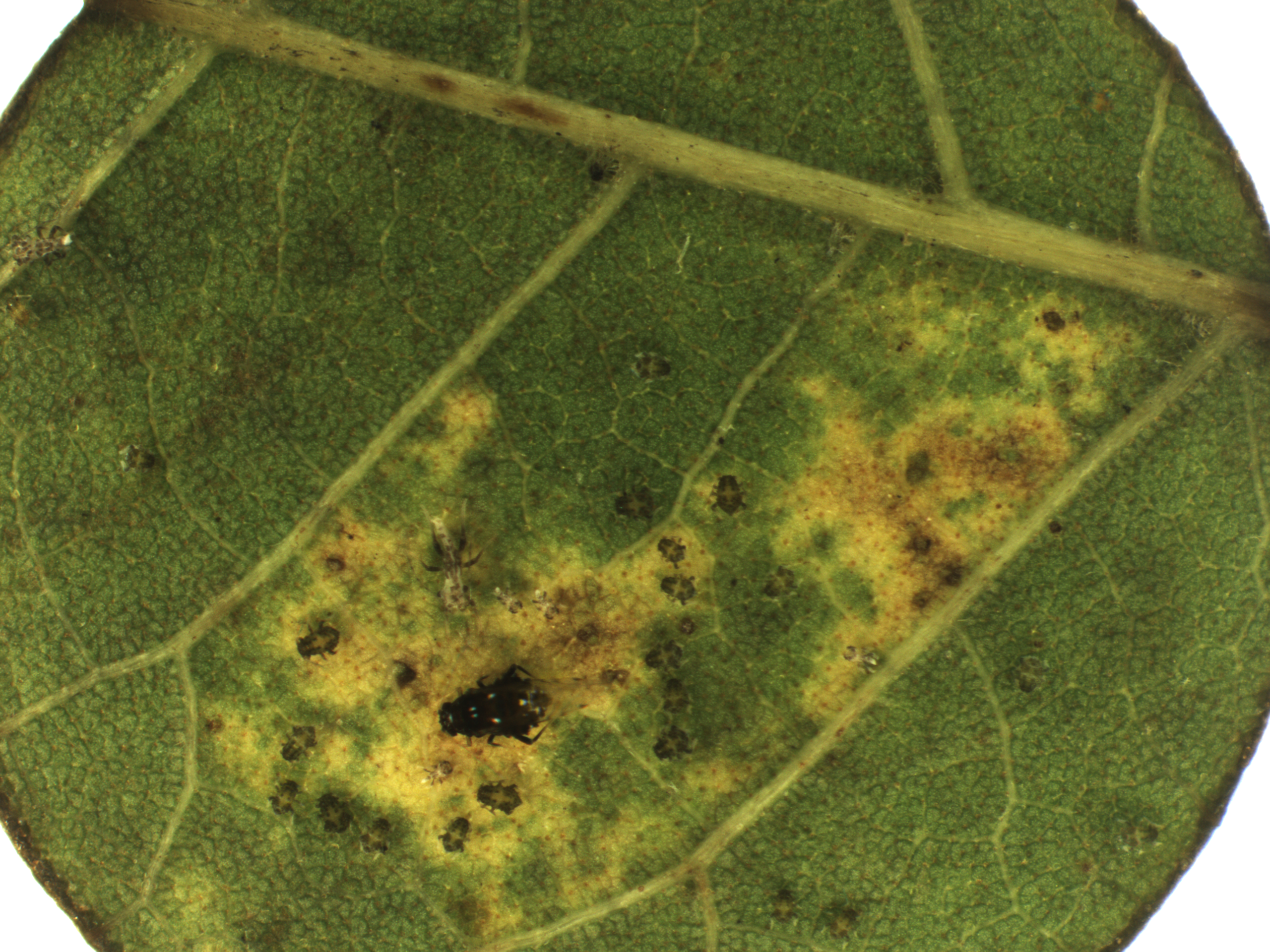
An adult black pecan aphid and her progeny. (Photo by Ted Cottrell)
By feeding on phloem sap, the black pecan aphid, blackmargined aphid, and yellow pecan aphid remove energy from the pecan tree. Although the physical process of inserting mouthparts into the leaf is similar for the three species, the blackmargined and yellow pecan aphids feed directly on phloem sap without any direct leaf modifications, whereas the black pecan aphid requires some modification to the leaf for feeding. Like many aphid species, the blackmargined and yellow pecan aphids rather quickly detect the suitability of the host plant, insert their mouthparts and feed. By feeding without leaf conditioning, the blackmargined and yellow pecan aphid nymphs can move from a feeding site when disturbed and begin feeding at a new site with what appears to be little impact on the aphid’s overall development, except for time lost moving from one site to the next. In contrast, the black pecan aphid takes a different approach to feeding and needs time to modify leaf tissue by breaking down leaf chlorophyll. Resulting leaf chlorosis is suspected from chlorophyll-degrading enzymes injected by the black pecan aphid.
Similarly, the Russian wheat aphid, while feeding on wheat, causes leaf chlorosis, and it is suspected that pectinases and cellulases are important salivary enzymes that lead to this chlorotic damage. It is likely that the black pecan aphid is feeding on the catabolites resulting from the enzymatic breakdown of chlorophyll. The black pecan aphid needs this process to elicit leaf chlorosis for feeding, but when the aphid withdraws early from a feeding site and begins anew, this process imparts a heavy penalty on the black pecan aphid due to the time required to break down the chlorophyll. Visible feeding damage by the black pecan aphid initially appears as a black dot where the leaf vein is punctured, followed by the surrounding laminar zone becoming chlorotic. When a newborn black pecan aphid begins feeding on a leaf, visible leaf chlorosis can show up within two to two and a half days. These chlorotic zones are typically bounded by leaf veins but can enlarge with the intensity of chlorosis being affected by physiological age of the leaf, duration of black pecan aphid feeding and age of the black pecan aphid nymph. During late summer and early fall, these chlorotic areas on leaflets dry out and turn brown as the leaf tissue dies.
Because the black pecan aphid relies on their associated chlorotic lesions for feeding, nymphs typically do not move far, if at all, from the initial feeding site through all molts to the adult stage. In a laboratory choice study using field-collected foliage, a higher number of black pecan aphids settled on pecan leaf discs that previously had been fed upon and damaged by the black pecan aphid than on leaf discs without chlorotic damage by the black pecan aphid. It was suggested in that study that the black pecan aphid was most likely attracted to, or arrested on, the damaged leaf discs due to the yellow coloration (chlorotic injury) imparted through earlier feeding injury by black pecan aphids. Many reports commonly state that aphids respond to certain colors. This statement does not discount the fact that volatiles could play a role in selecting damaged plant tissue and it is known that some aphids are influenced by volatiles.
When exposed to constant dark, adult black pecan aphids respond to chlorotic leaf discs rather than non-chlorotic leaf discs. This result seemingly negates visual attraction to chlorotic lesions and increases the possibility that volatile attractants are involved. However, aphids select a host through a sequence of steps in response to many stimuli and it is likely that random encounters with leaf discs by black pecan aphid adults under constant dark led to more aphids becoming arrested on chlorotic leaf discs. Under constant dark, more aphids probably chose chlorotic leaf discs that satisfied host selection criteria as they moved within the small Petri dish. This result contrasts with earlier reports stating that previous feeding injury by black pecan aphids reduced the performance of subsequent black pecan aphids feeding on the same foliage. This reduced performance was probably due to the duration and number of black pecan aphids feeding on foliage; leaf resources were likely exhausted and negatively affected subsequent feeding. Other reports state that by feeding, some aphid species can induce local changes in the plant that results in the development of larger aphids with higher reproductive capacity, which may account for black pecan aphids selecting previously damaged foliage.
Regardless of the factors affecting the black pecan aphid’s feeding site selection, elicitation of leaf chlorosis is important for normal black pecan aphid development. When nymphs of this species are allowed to feed continuously on the same leaf disc without interruption, they grow at a faster rate and resulting adults have a longer body than those adults arising from those nymphs not allowed to feed continuously on the same leaf disc (i.e., were not allowed to elicit chlorotic lesions on leaf discs). Leaves fed upon by black pecan aphids exude seven times the total amount of amino acids compared with non-infested leaves. An apparent preference for chlorotic leaves could be due to higher nitrogen availability within chlorotic lesions than within non-chlorotic leaves. Once the black pecan aphid initiates leaf chlorosis, the chlorotic area will increase in size even if the aphid is removed.

Development of a newborn black pecan aphid (nymph to the adult) showing how the aphid stays near the chlorotic lesion. (Photo by Ted Cottrell)
The black pecan aphid’s feeding practices cause this pest to straddle the line between two classifications for insect herbivores. Insect herbivores can be classified as flush feeders and senescence feeders according to the definitions for general nitrogen utilization by insect herbivores. Flush-feeding herbivores are adapted to survive on a relatively high concentration of readily assimilated nitrogen in young leaves for a brief period, while senescence feeders prefer tissues that have a relatively low amount of nitrogen over a longer period of time. Although the black pecan aphid is similar to senescence feeders regarding the foliage it feeds on, an increase in the abundance of amino acids in pecan foliage, after elicitation of chlorosis, makes this type of aphid similar to flush feeders, which utilize a higher concentration of nitrogen.
So why does a proportion of the black pecan aphid nymphal population move to the top surface of the pecan leaf? Some black pecan aphid nymphs make this move to have the time required to elicit chlorosis for feeding. Remember that many aphid predators spend most of their time searching the underside of the pecan leaf. If a black pecan aphid requires two to two and a half days to elicit chlorosis for feeding, then being disturbed and starting anew is almost certainly a death sentence due to the time required to elicit another chlorotic lesion and the increased exposure to predation over a longer period. In laboratory tests, black pecan aphid nymphs prevented from eliciting chlorotic injury had higher mortality than those nymphs allowed to elicit chlorotic feeding injury. For those black pecan aphid nymphs that did survive without feeding on leaf tissue that was visibly chlorotic (initial chlorophyll degradation may not be visible), it took them almost twice as long to develop to the adult stage and their body length was 40 percent shorter than adults that did elicit and feed on chlorotic zones.
Although these details of a black pecan aphid’s feeding habits are a lot to digest, this information is crucial to understand this pest fully. The final part of this series will cover how we can use this information regarding the black pecan aphid’s feeding biology as part of a management strategy. Part Three will also cover more information about the interaction of the black pecan aphid with the pecan tree and how certain plant bioregulators can negatively impact the ability of the black pecan aphid to elicit leaf chlorosis.